
 |
| A north African spinosaurine, with obvious nods to recent work suggesting some of these animals might've had short legs and a semi-aquatic lifestyle. The pterosaurs are azhdarchids, which are known to coexist with some African spinosaurines. |
The latest paper on these animals is that of Christophe Hendrickx et al. (2016), a piece which provides another interpretation on Moroccan spinosaurine diversity based on isolated quadrate bones. These are elements from the back of the skull which, among other things, articulate with the lower jaw. I don't really want to go into the ins and outs of their primarily descriptive and systematic assessment as the paper is a) a bit of a beast and b) we've spoken a lot about spinosaurine taxonomy of late and I'm desiring fresh topics. It will suffice to summarise that Hendrickx et al. (2016) provide compelling evidence for at least two spinosaurines being present in the Moroccan Kem Kem Beds, one of which is Sigilmassasaurus brevicollis and the other is - in their interpretation - Spinosaurus aegyptiacus. These results are not exactly the same as those presented in the recent Evers et al. (2015) paper, as Hendrickx et al. shuffle and deal north African spinsosaurid fossils among named taxa in another unique way. However, it certainly adds further evidence against the concept of a single spinosaurine species ruling Late Cretaceous north Africa proposed by Ibrahim et al. (2014). And yes, for those interested in scientific responses to the famous quadrupedal Spinosaurus reconstruction publicised in 2014, Hendrickx et al. (2016) specifically comment on the likelihood of it being a chimera of animals from across time and space. There's lots more in the paper - spinosaur skull morphology, body size, ontogeny, loads of illustrations and it's 100% open access - interested parties should definitely check it out.
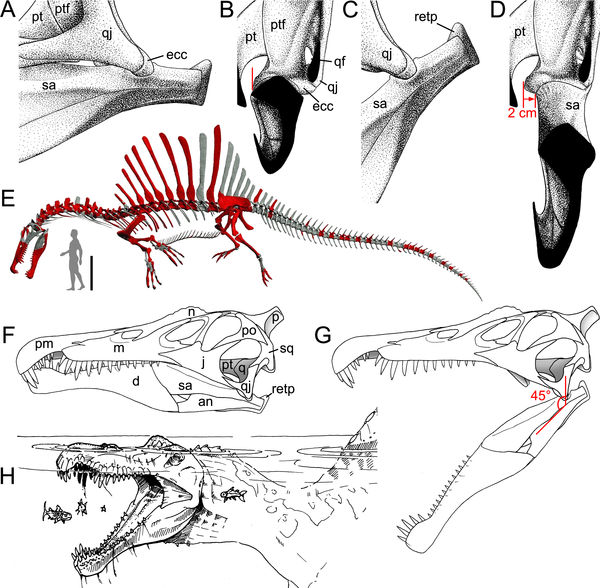 |
| The pelican-like foraging anatomy of Spinosaurus, as illustrated in Hendricks et al. (2016). |
*It's OK, my shame about that pun is worse than any punishment you could deliver.
In both the paper and news outlets (example), these perceived similarities between spinosaurid and pelican jaws are being stated as evidence of spinosaurs feeding in a pelican-like fashion, splaying their jaws to enhance food capture and swallow larger prey. Regular readers may recall that Darren Naish and I published research on similar claims for pterosaurs in 2013 (although the paper was not 'officially' published until last year - Witton and Naish 2015), countering suggestions that some pterosaurs fed like pelicans because of their helical quadrate articulations (Averianov 2013). This background made me surprised that another group of fossil animals was being labelled as pelican-like, as much of our discussion on pterosaurs is applicable to non-avian theropods and we're even cited in the Hendrickx et al. paper! I thought it might be of interest to explain some of my reservations about this idea here**.
**I want to note that I feel a bit awkward writing this commentary in light of recent controversies on palaeontology blogs, so-called 'Post Publication Peer Review' and so on. I hope that it's clear that this article, as with all the writings here, are meant as constructive, well-meaning expressions of opinion from someone with an interest in these topics and experience in a similar research field. My disagreement with the functional analogy proposed by Hendrickx et al. and their PR work is a polite one, and doesn't mean I disrespect them as scientists or want to undermine the significance of their paper.
We should begin by familiarising ourselves with the jaws of some relevant modern birds. Quite a bit of research has been done into pelican jaw anatomy (Schreiber et al. 1975; Meyers and Myers 2005; Field et al. 2011), an unsurprising fact given how awesomely and specifically adapted their jaws are to their unusual foraging method. Researchers have identified a number of adaptations critical to pelicans being able to splay their jaws so widely. They include reduced mineralisation at the middle part of the lower jaw to make a long 'bending zone', and even further reduction of mineral content (down to 20%) adjacent to the mandibular symphysis. This makes a 'hinge' for the mandibular rami to swing outwards on despite the fact there is no articulation or joint in the jaw at this point. The mandibular symphyses are so short that virtually all the jaw length is permitted to splay. At the other end of the jaw, the tongue is similarly reduced so as not to get in the way when the mouth is opened. The connection between the dentary bone (forming the jaw anterior) and the complex of bones of the posterior jaw is long, loosely connected and obliquely oriented so as to aid motion when the mandible spreads outwards. Even the horny tissues of the beak are specialised, being very thin (described as 'skin-like' by some authors) so as not to impede jaw flexion.
Contrastingly, little mention is made of helical jaw joints when talking about pelican foraging strategies. Zusi (1993) mentions that jaw spreading at the joints may be important to this effect, but more recent literature states that the precise mechanic responsible for bowing pelican mandibles is not really understood (Meyers and Myers 2005). Three hypotheses are currently thought viable: forces and weight of water acting on the jaw (known to be only part of the solution, as pelicans can splay their jaws on land too), contraction of muscles in the gular pouch (pulling the chin backwards, pushing the jaw rami out) or twisting of the lower jaw by the pterygoideus jaw musculature (Meyers and Myers 2005 and references therein). I guess it's possible that lateral displacement of the jaws has some role too, but it's interesting that the pelican guys aren't championing it's role as essential to manibular bowing. It's always risky using negative evidence in this way, but it might be telling given how intensively studied pelican skulls are.
 |
| Mandibular bowing in modern birds, as illustrated by Zusi (1993). a, herring gull (Larus argentatus) with relaxed and bowed mandibles; b, common potoo (Nyctibius griseus) skull and mandible (arrows show zones of flexion; c, tawny frogmouth (Podargus strigoides), a potoo incapable of mandibular kinesis. Note contrasting morphology between the potoo mandibles - even as a fossil taxon, there would be no doubt that N. griseus was capable of mandibular flexion. |
Modern birds give us a pretty good idea of what sort of features we're looking for in a pelican-like dinosaur. In doing so, as might already be obvious, they suggest major issues with the pelican-spinosaur analogy. Firstly, we need to acknowledge that helical jaw joints are not a special pelican feature. We could also call spinosaurid mandibular joints 'heron-like', 'potoo-like', or even 'hummingbird-like'. It seems more precise to suggest spinosaurid jaw joints are similar to those of several modern bird lineages and not over-emphasise anything to do with pelicans.
 |
| Our mandibular bowing experiments with pterosaurs and pelicans illustrated. Even when stretching the pterosaur jaws beyond the limits of their jaw joints, their area increase was negligible compared to that of a lazy pelican. From Witton and Naish 2015. |
My points here might be countered by the observation that the mandibular symphyses of some spinosaurids are quite fibrous, perhaps indicating a loose and mobile connection between them (Hendrickx et al. 2016). This might be the case, but Hendrickx et al. also note that some spinosaurid symphyses are longer than those of other theropods, thus actually having a greater degree of anterior attachment between each lower jaw. Based on our understanding of modern bird jaws, surely this is the opposite of what we'd expect in an extinct pelican-analogue? Even if I'm wrong on that, a slightly spongy symphysis and helical jaw joints are several functional miles off the flexibility afforded by avian jaws - especially those with the most extreme adaptations for mandibular bowing, like pelicans.
In all, then, I'm not convinced on the idea that spinosaurids were pelican-like in their foraging habits. Rather, the adaptations outlined by Hendrickx et al. suggest a somewhat bird-like ability to increase gape for swallowing slightly larger food than usual. In actuality, spinosaurid jaw functionality contrasts so markedly with that of pelicans that it might be misleading to tout pelicans as spinosaur analogues. After all, the spinosaurid ability to bulge their jaws slightly is not especially pronounced or even rare, whereas what pelicans do is really both those things: an extreme and marked adaptation, and almost unique in nature. I come back to a point I've made about other instances of 'extreme' modern animals being used as analogues for extinct ones: we need to be as thorough as possible in our functional assessments before pointing to highly specialised and extremely adapted modern species as suitable analogues for long dead taxa.
Enjoyed this post? Support me on Pateon!
The artwork and words you see here are supported by Patreon, the service which allows you to directly support artists and authors with monthly payments. You can support this blog from $1 a month and, in exchange, get access to bonus content and rewards - the more you pledge, you more bonuses you receive. For this post, I'll be explaining the rationale behind the spinosaurine depicted at the top of this post - it's not Spinosaurus 2014, not a 'classic' reconstruction - what's going on?
Later this month (February 2016) I'll be sending out signed art prints to all those who pledge at least $5 a month - sign up now to get yourself on the receivership list!
Later this month (February 2016) I'll be sending out signed art prints to all those who pledge at least $5 a month - sign up now to get yourself on the receivership list!
References
- Averianov, A. O. (2013). Reconstruction of the neck of Azhdarcho lancicollis and lifestyle of azhdarchids (Pterosauria, Azhdarchidae). Paleontological Journal, 47(2), 203-209.
- Field, D. J., Lin, S. C., Ben‐Zvi, M., Goldbogen, J. A., & Shadwick, R. E. (2011). Convergent evolution driven by similar feeding mechanics in balaenopterid whales and pelicans. The Anatomical Record, 294(8), 1273-1282.
- Hendrickx, C., Mateus, O., & Buffetaut, E. (2016). Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa. PloS one, 11(1), e0144695.
- Ibrahim, N., Sereno, P. C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D. M., Zouhri, S. Myhrvold, N. & Iurino, D. A. (2014). Semiaquatic adaptations in a giant predatory dinosaur. Science, 1258750.
- Louchart, A., Tourment, N., & Carrier, J. (2011). The earliest known pelican reveals 30 million years of evolutionary stasis in beak morphology. Journal of Ornithology, 152(1), 15-20.
- Meyers, R. A., & Myers, R. P. (2005). Mandibular bowing and mineralization in brown pelicans. The Condor, 107(2), 445-449.
- Schreiber, R. W., Woolfenden, G. E. & Curtsinger, W. E. (1975). Prey capture by the Brown Pelican. The Auk, 92(4), 649-654.
- Witton, M. P. and Naish, D. (2015) Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"? Acta Palaeontologica Polonica 60, 651-660.
- Zusi, R. L. (1993). Patterns of diversity in the avian skull. The skull, 2, 391-437.
First. Sorry :(
ReplyDeleteThe authors do compare the helical mandibular joint to pelicans, pterosaurs, and other fast biting/weak jawed theropods I think that they do this more on the presumption of piscivory and/or swallowing prey whole and looking for a suite of characteristics that unite this lifestyle. Yes they do compare spinosaurids and pelicans but - as you mentioned - the unique bowing of the mandible for pelicans really is what sets them apart. From what I gather reading their paper it is the combination of loosely connected mandibular symphysis and the helical joint that allowed deglutition of large food parcels in spinosaurids. Really I think it is the press that spun this comparison between pelicans and spinosaurs to an extreme. The question that needs asking is how far the mandibular symphysis could spread apart - maybe snakes offer some comparison?
ReplyDeleteNow there's an interesting idea. I, too, felt that the pelican comparison was a tad too extreme.
DeleteHi Mark,
ReplyDeleteThanks for your comments and interest in my research, appreciated! I just wanted to say that we basically share the same opinion about similarities and differences between spinosaurs and pelicans, and that the large majority of your comments do not really go against what is written in this papers. We mostly compare the jaw mechanism of spinosaurs with pelicans mainly because it was done by other authors in regards to the diagonally oriented helicoidal intercondylar sulcus of the mandibular articulation of the quadrate, also seen in pterosaurs. And, because spinosaurs have a mandibular symphysis restricted to the anterior part of the jaw and linked by connective tissue, we made a further comparison with pelicans (as the pterosaur mandibular symphysis extends much along the dentary and is fused as far as I know). That's pretty much it and we are deeply aware, my colleague and I, that the mandibular ramus of spinosaurids and possibly most non-avian theropods could not bent like that of pelicans and that spinosaurids probably did not have a pouch (which is why we didn't add one in the reconstruction of the two spinosaurids by Sergey Krasovskyi). Likewise, we are aware that the lateral movement of the two rami was not that significant, yet you often refer to the length of the skull when talking about this but the most important aspect of this lateral displacement goes with the width of the back of the skull, which is relatively narrow in spinosaurids, and might have been even narrower in derived spinosaurs. And when you say "we could also call spinosaurid mandibular joints 'heron-like', 'potoo-like', or even 'hummingbird-like'", I totally agree with you. Yet people tend to portray this wideness of the mouth much better when we tell them it's similar to that seen in pelicans. And I don't really think we are misleading them by saying that. Likewise, because this adaptation in spinosaurs helped them to swallow large prey items a whole, and most likely fish considering they were at least partially piscivorous animals, the comparison with pelicans is not stupid either. Hope you agree with my points, Duane Nash qt least seems to share my opinion on this regards! Cheers, C.
Hi guys,
ReplyDeleteThanks for the comments. Perhaps the issue here is that the 'established' go-to bird for lateral motions at jaw joints is now known to have a lot of functional baggage that complicates it as a suitable analogue for most other animals (as noted by some recent research, the only animals with pelican-like jaws are rorquals). Given what research conducted in the last few years has shown, maybe it's time to move away from pelicans - or at least include other birds - when talking about jaw motion. Christophe, as you indicate, this can be more complex when dealing with the public as the pelican is the best known bird for this trick, but one neat thing about dinosaur science is that it provides a gateway to introducing new concepts to lay audiences. In this instance, spinosaurs can be used to introduce the fact that a lot of birds have mobile jaw joints - pelicans are just one (and an extreme) example.
Yet another paper criticizing Ibrahim's reconstruction. This is why I wasn't quick to jump on the bandwagon and get caught up in all the height behind the new findings. It's best to wait a few years and see what new evidence and subsequent analysis of the material before jumping to conclusions. Still, all this news about Spinosaurus is really exciting. After so many years of almost nothing new being proposed about this fascinating creature, the 2014 find and the subsequent discoveries made in recent years is really exciting! Spinosaurus has always been a favorite of mine, and all these new finds and theories are delightfully thrilling! Thanks for posting this, Mark, especially that link to the new paper!
ReplyDeleteSo out of curiosity I went around on the internet and looked at some other blogs regarding the spino's life appearance and found this:
ReplyDeletehttp://antediluviansalad.blogspot.com/2016/03/spinosaurus-unauthorized-iii-run-spino.html
I'm not sure if you have read it already but I thought the concept wasn't too far-fetched, maybe. I think it's a three part post. Just a thought.
This doesn't look good for my theory that Spinosaurus would run toward the water, launch itself over the water with its mandible in the water to skim feed while simultaneously storing the caught prey in a pelican-like gular pouch.
ReplyDeleteI had somehow whiffed on this post when it first came out, so I'm glad it got linked somewhere (FB I think). Interesting stuff both in a number of dimensions - extant/extinct - birds/other theropods - spinosaurs/pterosaurs.
ReplyDelete